Лимфоидная ткань представляет собой систему лимфоцитов и макрофагов, которые находятся в клеточно-волокнистой ретикулярной строме. Эти клетки функционируют в составе либо обособленных органов, либо диффузных образований. Лимфоидная ткань является функционирующей паренхимой лимфоидных органов - костного мозга, вилочковой железы, селезенки, лимфатических узлов, лимфоидной ткани кишечника и соединительной ткани. Участки лимфоидной ткани находятся в слизистой оболочке некоторых органов (бронхов, мочевых путей, почек). Для профилактики пейте Трансфер Фактор . Лимфоидные органы относятся либо к первичным, либо к вторичным органам. Первичные лимфоидные органы - это красный костный мозг и вилочковая железа. Первичные лимфоидные органы служат основным местом развития лимфоцитов. К вторичным лимфоидным органам относятся лимфатические узлы, селезенка и скопления диффузной лимфоидной ткани в слизистых оболочках дыхательных, пищеварительных и мочеполовых путей. Вторичные лимфоидные ткани заселены клетками ретикулярного происхождения, а также макрофагами и лимфоцитами, предшественниками которых служат стволовые клетки костного мозга.
Лимфоидная ткань принимает самое активное участие во всех без исключения защитных реакциях организма. Она рассеяна практически по всему организму и контролирует определённые его участки. Основу лимфоидной ткани составляют ретикулярные волокна и ретикулярные клетки, которые формируют сеть с ячейками различной величины. В петлях сети находятся клетки лимфоидного ряда, среди которых малые, средние и большие лимфоциты, плазматические клетки, молодые клетки - бласты, макрофаги, а также небольшое количество лейкоцитов, тучных клеток. Ретикулярная строма образуется из мезенхимы, а клетки лимфоидного ряда — из стволовых клеток костного мозга, обеспечивающего кроветворение (миелопоэз) и формирование клеток иммунной системы (лимфопоэз). Среди органов и тканей этой системы имеются истинно лимфоидные образования, в которых происходит только лимфопоэз - это вилочковая железа, лимфатические узлы, лимфоидная ткань кишечника и "смешанные" образования, где находятся как лимфо-, так и миелопоэз - костный мозг, селезенка. Клетки лимфоидного ряда, среди которых выделяют две популяции - Т- и В-лимфоциты, перемещаются с кровью и лимфой. Вместе с макрофагами они участвуют в реакциях иммунного ответа против генетически чужеродных веществ. Если через ссадину, рану попадает чужеродный антиген, первым вступает в защитную реакцию ближайший к месту проникновения лимфатический узел. Если антигенная диверсия массивна и не подавляется местными силами - в защиту вовлекается вся система иммунитета.
Лимфоидная ткань человека составляет около 1% веса тела и является одним из важнейших компонентов лимфоидных органов. Эта ткань очень чувствительна к внешним и внутренним воздействиям. Так, под действием рентгеновских лучей лимфоциты быстро погибают. При введении тиреоидных гормонов лимфоидная ткань разрастается. Большое влияние на степень развития лимфоидной ткани имеют гормоны коры надпочечников. Недостаточная функция коры надпочечников вызывает разрастание лимфоидной ткани. Введение гормонов коры надпочечников приводит к дегенерации лимфоидной ткани и гибели лимфоцитов. По мере старения организма уменьшается количество лимфоидной ткани и лимфоидных узелков в органах иммунной системы. При воспалительных процессах и активации иммунных реакций, как первичного, так и вторичного характера, наблюдается реактивная гиперплазия лимфатических узлов.
Параллельно велись исследования другой системы организма, напрямую не связанной с лактацией, но тем не менее играющую колоссальную роль в защите внутренней среды организма от внешних воздействий. Исследование кишечника, а более конкретно - лимфоидной ткани желудочно-кишечного тракта ведется достаточно давно: с середины прошлого века в лимфоидной ткани кишечника выделяли такие компоненты, как кишечные лимфожелезистые комплексы.
Это округлые скопления (диаметром 0,5-5 ми) трубчатых ветвящихся желез, или крипт, окруженные лимфоидной тканью. Их общее количество во всей ободочной кишке было 325-791, причем в первом сегменте их количество колебалось от 2 до 15, во II пятой части - 31-216, в III пятой части - 17-243, в IV пятой части - 122-155 и в последней (задней) пятой части - 62-215. Авторы обнаруживали их макроскопически, они видны при просвечивании кишечной стенки, особенно после снятия серозной оболочки. По диаметру и клеточному составу (главные и бокаловидные энтеро- циты) эти железы были сходны с кишечными криптами, и, пробуравливая мышечный слой слизистой, они открывались либо в кишечные железы, либо самостоятельно на поверхность слизистой оболочки. Располагались такие железистые скопления субмукозы в кишечной стенке без определенного порядка.
Предполагается, что они образуются из зачатков обыкновенных кишечных желез путем впячивания последних в подлежащую подслизистую оболочку в утробном периоде.
Агрегированные лимфатические узелки тонкого кишечника встречаются в тощей и подвздошной кишках всех домашних животных, а у собаки, кроме того, и в дуоденуме. Обычное место их локализации - антимезентериальная стенка кишечника, а у собаки и кошки наряду с этим и вблизи брыжеечного края кишки. Некоторая асимметрия в виде правостороннего парамедиального расположения агрегированных узелков наблюдается в каудальной части тонкой кишки свиньи.
Это явление объясняется некоторым отклонением от вертикального расположения брыжейки и кишечника. Полагают, что в организме живой свиньи правая сторона кишечной стенки имеет право-вентральное расположение, и что вследствие этого в случае неполного наполнения кишечной трубки большая часть всасывающей поверхности образуется правой половиной кишечной стенки. Здесь же всасываются и более многочисленные патогенные агенты, попадающие в лимфатические узелки.
В слизистой толстой кишки встречаются многочисленные одиночные лимфатические узелки; агрегированные же узелки отмечаются редко, ограничиваясь при этом у крупного рогатого скота и лошади верхушкой слепой кишки и у жвачных и свиньи - окружностью илеоцекального отверстия. У жвачных, кроме того, находится постоянно одна узелковая пластинка в конце начального завитка ободочной кишки. У лошади более тесное скопление лимфатических узелков обычно отмечается также в области тазового изгиба большой ободочной кишки. На месте узелков слизистая либо слегка приподнята, либо углублена; в углублениях железы обычно отсутствуют. Лимфоидная ткань толстой кишки наиболее развита у свиньи.
Большое количество лимфоидных клеток встречается в ретикулярной ткани. Она очень богата различными клеточными элементами - фибробластами, плазмоцитами, макрофагами, лимфоцитами, эозинофилами и недифференцированными клетками.
Совокупность названных клеток совместно с межклеточным веществом стромы составляет мощный барьер для различных микроорганизмов и любых антигенных веществ содержимого кишечника, на что, впрочем, указывает и значительно менее развитая клеточная популяция у стерильных животных. В наибольшем количестве встречаются фибробласты (в кишечнике мыши их из всего клеточного состава стромы слизистой в среднем 45,57%); в виде уплощенных клеток они образуют под базальной мембраной непрерывный слой. Плазматические клетки находятся весьма часто в контакте с макрофагами, от которых они, вероятно, получают необходимую информацию для окончательной дифференциации. Лимфоциты чаще всего располагаются близ кровеносных капилляров. Вблизи сосудов располагаются и лабро- циты, которые, однако, в большем количестве многочисленно встречаются в субмукозе. Часть гранулоцитов и лимфоцитов подвергается беспрерывной эмиграции через эпителиальный покров, особенно в тонком кишечнике. В содержимом кишечника они разрушаются, выделяя при этом бактерицидные вещества, оказывающие избирательное действие на естественную микрофлору кишечника. Общее количество тех или иных клеток зависит от вида животного, его возраста, от функционального состояния органа и т. д.
Меж- и подкриптальная строма слизистой всюду пронизана сетью аргирофильных волокон. Эластические и (особенно) коллагеновые волокна встречаются в незначительном количестве. Отмечаются в межкриптальной строме и некоторые гладкомышечные клетки, а в ворсинках они являются обычными структурными и функциональными элементами. Эластические и коллагеновые волокна сопровождают в первую очередь кровеносные сосуды, а сеть аргирофильных волокон в более сгущенном виде окружает кишечные крипты. Эластические волокна более развиты у плотоядных животных.
Из наиболее изученных животных самое большое количество свободных клеточных элементов слизистой пищеварительного тракта наблюдается у свиньи, а самое маленькое - у лошади и плотоядных. Наиболее богатой свободными клетками является тонкая кишка. В тонкой кишке показатели количества плазматических клеток и лимфоцитов взаимно противоположны. На уровне кишечных крипт количество плазматических клеток часто достигает 100 и более, но в ворсинках в несколько раз уменьшается, а количество лимфоцитов между криптами, наоборот, менее значительное, но повышается в ворсинках. Количество блуждающих клеток тесно связано с характером желез данной области и с их функциональным состоянием. Это особенно наглядно обнаруживается при сравнении количества свободных клеток между дуоденальными криптами и соединительной тканью дуоденальных желез. С другой стороны, количество мигрирующих клеток весьма тесно связано с характером корма данного вида животного.
У новорожденного ягненка в собственном слое слизистой встречаются только единичные лимфоциты и гранулоциты; плазматические клетки появляются после рождения, и они достигают своего максимума (около 120 клеток в поле зрения микроскопа при 600-кратном увеличении) к 5-6-месячному возрасту, а лимфоциты в 2-летнем возрасте, причем их число достигает 400 и более. У взрослой овцы в наибольшем и наиболее постоянном количестве встречаются лимфоциты; за ними следуют плазматоциты, наибольшее число которых отмечается в дуоденуме. Во всей пищеварительной системе самым высоким является общее количество свободных клеток в lamina propria мукозы тонкого кишечника. Количество гранулоцитов колеблется в значительной степени, причем эти колебания зависят не от возраста овцы, а от функционального состояния данного органа.
Кроме этого, указывается на большое количество рассеянных лимфоидных клеток, так называемых глобулярных лейкоцитов - округлых внутриэпителиальных клеток, содержащих в своей цитоплазме шаровидные оксифильные зернышки (глобулы). Они встречаются как в покровном, так и в железистом (криптальном) эпителии всего желудочно- кишечного тракта позвоночных (Kent, 1949); описаны они также в трахее, мочевом пузыре и в половых органах. Впервые глобулярные лейкоциты были описаны в 1920 г. Вейлом. Их почти постоянная локализация в эпителиальной ткани оправдывает их описание рядом с различными видами эпителиальных клеток кишечника.
На мощное развитие лимфоидной системы указывалось достаточно давно. Лимфатическое русло кишечной стенки состоит из взаимно связанных собственно-слизистого, подслизистого, межмышечного и подсерозного сплетений лимфатических капилляров и сосудов. Эти сплетения более детально изучены у кошки в тонком кишечнике и в толстой кишке. Разделение интрамурального лимфатического русла на лимфатическую сеть, образованную лимфатическими капиллярами, и сплетения, формируемые лимфатическими сосудами, у кошки выражены нерезко. Эта сеть состоит (в толстой кишке) из полигональных петель, образованных из лимфатических сосудов, содержащих клапаны, и капилляров. При этом стволики слепо начинающихся коротких лимфатических капилляров могут впадать непосредственно в лимфатические сосуды, не образуя капиллярной сети.
В тонком кишечнике лимфатическая сеть мукозы располагается между криптами и у корней ворсинок. В последние открываются центральные млечные сосуды - своеобразные лимфатические капилляры шириной от 5 до 10 мкм и длиной до 600 мкм. В подслизистой оболочке залегают три вида капиллярных лимфатических сетей, тесно связанных друг с другом и образованных капиллярами различного диаметра. Мелкие петли, образованные капиллярами, обрамляются петлями крупнопетлистой сети. Ближе к брыжеечному краю из капилляров крупнопетлистой сети формируются собирательные лимфатические сосуды, которые прободают циркулярный мышечный слой и сливаются с собирательными лимфатическими сосудами межмышечного сплетения или самостоятельно поступают в толщу брыжейки. В межмышечном сплетении внутри крупнопетлистой сети (состоящей из сосудов диаметром 30-45 мкм) лежат мелкие петли из более тонких (15-30 мкм) капилляров.
Собирательные и отводящие лимфатические сосуды кишечной стенки снабжены клапанами. Каждый из лимфатических узелков также окружен сетью лимфатических капилляров.
Утверждение некоторых других авторов о том, что млечные сосуды ворсинок являются артефактами, в естественных условиях не существующими лимфатическими капиллярами, не соответствует результатам исследований большинства современных авторов. Млечные капилляры ворсинок выстланы плоским эпителием, окутанным снаружи ретикулярными волокнами и гладкомышечными пучками. Микрокапли жира переносятся в просвет капилляра с помощью пинодитарного транспорта. Среди факторов, воздействующих на ток хилуса, абсорбированного из кишки, видную роль играют ослабление и сокращение мышечного слоя слизистой оболочки кишечника. В стенке тонкой кишки кошки и собаки различают две различных системы лимфатических сосудов: одна отводит лимфу в состоянии покоя, а другая включается лишь в случае повышенного образования лимфы.
Однако до недавнего времени все-таки оставалось не выясненным, проникают ли клетки молозива через кишечную стенку или нет и если да, то каким образом. Ответ на этот вопрос был получен в 2002 г. Tuboly S., Nelson D. L. изучали кишечное поглощение молозивных лимфоидных клеток.
Опыт проводился на 23 поросятах (от 4 свиноматок) и 17 ягнятах. От молозива и крови маток лимфоидные клетки были изолированы с помощью Фиколл-Пака и маркированы технецием. Через 7 часов после рождения клеточные суспензии объемом 10 мл каждая (поросята: 10 (7) клеток, ягнята: 5 х 10 (7) клеток) были введены после лапаротомии непосредственно в желудок или в тощую кишку через носоглоточный зонд. Замороженные срезы двенадцатиперстной кишки, тощей кишки и образцов лимфатического узла животных были исследованы авторадиографией.
Было обнаружено, что лимфоидные клетки, присутствующие в молозиве поросенка и ягненка, от их собственной матери были поглощены через пищеварительный тракт и через лимфатические сосуды транспортировались к брыжеечным лимфатическим узлам. Электронная микроскопия показала, что поглощение происходит межклеточно. Моло- зивные клетки от другой матки (не собственная мать поросенка), т. е. аллогенные клетки, лимфоидные клетки, изолированные от крови, и пастеризованные молозивные лимфоидные клетки не были поглощены. Иммунизация овец и их ягнят анатоксином столбняка демонстрирует, что поглощенные лимфоидные клетки остаются иммунологически активными, и могут передавать иммунную информацию к ягнятам.
Итак, во всяком случае для двух видов млекопитающих передача иммунной информации посредством клеточных компонентов доказана. Кишечная стенка, таким образом, представляет собой своеобразный контрольно-пропускной пункт со своей сложной системой регуляции и частично автономной системой иммунных органов.
Если обратиться к более ранним источникам, то можно узнать, что лимфатическая система детеныша проводит своеобразную подготовку лимфоидной ткани к приему иммунной информации. Во всяком случае об этом может говорить давно установленный факт: у поздних плодов в слизистой желудочно-кишечного тракта появляются зернистые формы лейкоцитов: эозинофилы и нейтрофилы. Они раньше появляются и в большом количестве концентрируются в тонком отделе кишечника, особенно в подвздошной кишке. Здесь же у поздних плодов овец и свиней появляются глыбчатые лейкоциты. Их образование начинается с проникновения лимфоцитов из соединительной ткани слизистой в пространства между эпителиальными клетками основания ворсинок и крипт. У таких лимфоцитов протоплазма теряет базофильные свойства и начинает вырабатывать специфические эозинофильные гранулы, значительно увеличивающиеся в размерах. Гранулы часто свободно лежат между эпителиальными клетками. Тогда же было обнаружено и явление ухода лимфоцитов из межэпителиального пространства в соединительную ткань слизистой. Плазматические же клетки появляются в слизистой кишечника овец и свиней до рождения, а у крупного рогатого скота - после рождения, что связано, очевидно, с поступлением молозива, содержащего значительное количество иммуноглобулинов.
Таким образом, для некоторых сельскохозяйственных животных вопрос передачи иммунитета посредством лейкоцитов молозива оказался более или менее выясненным. Что касается других двух видов домашних животных, то тут гораздо меньше информации имеется по механизмам передачи иммунитета посредством молозива у хищных домашних животных. Несмотря на то, что они относятся к млекопитающим, у них могут оказаться какие-либо видовые особенности.
В вопросах передачи иммунитета нельзя забывать о других путях транспортировки иммуноактивного материала, например, о трансплацентарном. Давно известно о наличии разных видов плацент и о их различной проницаемости для иммунноактивных компонентов. Однако подробно корреляция между клеточным составом молозива, его цитопрофилем и типом плаценты, характерной для этого животного, практически не обсуждалась. Достаточно четкая связь между составом первых порций молозива и продолжительностью периода лактации рассмотрена нами у приматов (Скопичев В. Г., Гайдуков С. Н., 1991). Конечно, прогнозирование продолжительности периода лактации у мелких хищников, таких как собаки и кошки, значительной хозяйственной необходимости не имеет, однако подобные данные могли бы оказаться весьма полезными при промышленном получении молока, например, от коров.
Согласно современным представлениям иммунная система как неотъемлемая часть организма участвует в подготовке молочной железы к периоду лактации. Причем участие клеточных элементов иммунной системы в формировании молочной железы настолько велико, что подавление деятельности регионарного лимфатического узла (источника лимфоидных клеток) существенно нарушает морфологические процессы и подготавливает развитие структуры альвеолы. В молозивный период в молочной железе наблюдается значительное увеличение клона Т-лимфоцитов и плазматических клеток. Причем рост количества плазматических клеток может наблюдаться у ряда животных и в течение молочного периода (Скопичев В. Г., Гайдуков С. Н., 1991). Кроме того, наблюдается увеличение количества фракций нейтрофилов, моноцитов. В момент проведения нами исследований были обнаружены делящиеся лимфоциты, что способствует увеличению численности их фракций.
С момента выделения с молозивом клеточные элементы и иммуноглобулины попадают в желудочно-кишечный тракт новорожденного и включаются в его механизмы формирования иммунной защиты. Временной диапазон передачи материнских антител млекопитающих различен. И по этому признаку их можно разделить на три большие группы: а) послеродовая передача материнских антител, сюда можно отнести всех копытных (жвачных, лошадей и свиней); б) группа с предродовой передачей антител - морская свинка, кролик, человек; в) группа животных, обладающих пред- и преимущественно послеродовой передачей - собака, мышь и крыса. У собак и мышевидных грызунов передача антител продолжается в молочный период лактации. Материнские антитела могут попадать в кровообращение плода до рождения через сосуды желточного мешка или плаценту, а после рождения - с молозивом через стенку кишечника и служить ему, как и взрослым особям, для защиты против внедряющихся в организм возбудителей. Пассивная передача материнских антител плодам в утробный период во многом определяется типом плаценты.
Эпителиохориальная (у моногастричных копытных) и синдесмохориальная (у жвачных животных) плаценты состоят из шести слоев и не пропускают антитела; эндотелиохориальная плацента (у плотоядных) состоит из четырех слоев, и антитела проходят через нее ограниченно; гемохориальная плацента (у приматов, грызунов) устроена из трех слоев и пропускает полностью или частично антитела; гемоэндотелиальная плацента (у морских свинок, кроликов на поздней стадии беременности) имеет один слой, и через нее полностью проходят антитела матери. Однако и здесь проницаемость плаценты для разных классов иммуноглобулинов различна. Иммуноглобулины классов IgA, IgE и IgM практически не проходят через плаценту, а из иммуноглобулинов класса G фракция IgGi проходит лучше, чем IgG 2 .
Плацента и сама вырабатывает вещества, регулирующие отношения матери и плода, в том числе гормоны (хо- риотропинальный соматотропин, гонадотропин, лютотропин, тиротропин, кортикотропин, эстрогены, прогестероны и др.) Кроме того, плацента синтезирует и воспринимает огромное количество ростостимулирующих факторов (интерлейкин-1, интерлейкин-2, КСФ) и ростоингибирующими (ФНО-а, ТРФ-Р, интерлейкин-б, интерферон). При этом ФНО-а повышает резистентность к ЕК-клеткам, ИЛ-6 активирует клетки супрессоры (Т-супрессоры). Интерфероны (ИФ-а и ИФ-р), блокируя процессы пролиферации эффекторных лимфоцитов, защищают плод от иммунного цитолиза. Известно, что плацента блокирует проникновение антител матери в кровеносное русло плода. Ранее мы отмечали, что в плаценте были обнаружены специфические F-рецепторы почти к всем четырем классам иммуноглобулинов. Именно поэтому у большинства животных не наблюдается гемолитической болезни. Однако с другой стороны, именно в этой ситуации возрастает роль послеродовой передачи иммунитета. Ранее авторами были сделаны попытки обнаружить факты проникновения клеток из молозива в организм детеныша. Для этих целей было решено использовать естественную метку клеток самок - половой хроматин. Метод основан на исследовании структурного образования в ядрах клеток - Х-хроматина. Интенсивность окраски Х-хроматина выше, чем других участков хроматина ядра; тельце Х-хроматина, кроме того, выделяется при любой его форме четкими контурами.
При этом методе в ядерных клетках у самок обнаруживается включение - половой хроматин, в клетках самцов он не обнаруживается. Для исключения лейкоцитов детенышей- самок в качестве подопытных были оставлены только детеныши-самцы. После выпаивания молозива проводилось изъятие крови и из внутренних органов (селезенка, лимфатические узлы, печень) приготавливалась лейкоцитарная взвесь, которая и шла в дальнейшее исследование. Далее нашей задачей были оценка свойств молозивных лейкоцитов травоядных и плотоядных животных и оценка их влияния на клеточный иммунитет у этих разных групп животных.
Функциональное состояние лейкоцитов могло, по нашему мнению, оказаться одним из ключей к разгадке их роли в трансколостральной передаче иммунитета. Влияние молозива на иммунную систему потомства очень хорошо изучено для гуморальных факторов, таких как иммуноглобулины и немного хуже для клеточного компонента. Однако исследования иммунного статуса были все-таки включены для полноты картины изменений в клеточном иммунитете детенышей в течение молозивного периода.
В процессе передачи иммунитета большое значение имеет и состояние иммунной системы реципиента. В нашем случае исходя из данных, представленных в обзоре литературы, большое значение необходимо придавать лимфоидной ткани (MALT), связанной со слизистыми, и в частности, лимфоидной ткани (GALT), связанной с кишечником, как регионарному представителю иммунной системы и ее форпосту на пути молозивных клеток. Для более детального изучения лимфоидной ткани, связанной с кишечником, было проведено гистологическое исследование кишечника кошек (котят) до рождения и в течение молозивного периода.
Проводилось выделение лимфоцитов из крови и молозива с целью исследования их биохимических и иммунологических свойств. Оценка фагоцитоза in vitro производится в соответствии с фазами реакции: через 30 минут и 2 часа. Поглотительная способность клеток оценивается по двум показателям: процент фагоцитоза, количество фагоцитов на 100 нейтрофилов через 30 минут и через 2 часа инкубации (в процентах), ФИ - среднее число микробов на 1 фагоцит через 30 минут и 2 часа инкубации. О последней фазе фагоцитоза - переваривании - судят по коэффициентам процента фагоцитоза и ФИ (отношение соответствующих показателей, изученных через 2 часа контакта, к тем же показателям через 30 минут). Фагоцитоз считается завершенным при коэффициентах менее 1.
Иоффи и Куртис (Yoffey, Courtice, 1970) объединили лимфоидную и кроветворную системы в единый лимфомиелоидный комплекс (рис. В.6).
Комплекс представляет собой систему органов и тканей, паренхима которых содержит клетки мезенхимального происхождения. В него входят: костный мозг, тимус, селезенка, лимфатические узлы, лимфоидная ткань кишечника и соединительная ткань.
Функциональные клетки лимфоидной системы представлены лимфоцитами, макрофагами, антигенпрезентирующими клетками и в некоторых тканях эпителиальными клетками. Все эти клетки функционируют в составе либо обособленных органов, либо диффузных образований.
Лимфоидные органы относят либо к первичным (центральным), либо ко вторичным органам. Первичные лимфоидные органы - это красный костный мозг и тимус.
Функциональное назначение комплекса - обеспечение кроветворения (миелопоэза) и формирование клеток иммунной системы (лимфопоэза). Среди органов и тканей комплекса имеются истинно лимфоидные образования, в которых происходит только лимфопоэз (тимус, лимфатические узлы, лимфоидная ткань кишечника) и "смешанные" образования, где представлены как лимфо-, так и миелопоэз (костный мозг, селезенка).
Именно в первичных органах формируется репертуар специфичностей лимфоцитарных антигенраспознающих рецепторов, и лимфоциты приобретают таким образом способность распознавать любые антигены, с которыми организм может столкнуться в течение жизни. Далее эти клетки подвергаются отбору на толерантность (ареактивность) к аутоантигенам, после чего уже в периферических лимфоидных органах или образованиях распознают только чужеродные антигены.
В тимусе, кроме того, T-клетки "учатся" распознавать собственные молекулы MHC . Вместе с тем известно, что некоторые лимфоциты развиваются вне первичных органов.
Из первичных органов лимфоциты мигрируют для выполнения своих функций по кровеносному руслу в периферическую лимфоидную ткань - лимфатические узлы, селезенку и лимфоидную ткань слизистых оболочек (пейеровы бляшки, миндалины). Это движение лимфоцитов от центральных органов иммунной системы на периферию является главным миграционным путем. Кроме того имеется путь рециркуляции. Лимфатические сосуды, дренирующие тело, собирают внеклеточную жидкость - лимфу - вместе с рассеянными по телу лимфоцитами и переносят ее в лимфатические узлы. После некоторого времени пребывания в лимфатических узлах лимфоциты собираются в выносящих эфферентных лимфатических сосудах. Из них лимфоциты попадают в основной лимфатический сосуд - грудной проток, откуда вновь возвращаются в кровоток через левую подключичную вену (рис. 6.1 и рис. 6.2).
Таким образом, лимфоциты относятся к той категории клеток, которые широко распространены в организме. И в теле человека и позвоночных животных они сгруппированы в три типа объединений (рис. 6.14). Различные типы организации лимфоцитов обеспечивают наиболее эффективное проявление лимфоидной системы при встрече с чужеродным антигеном.
Иммунный ответ на антигены, поступающие в организм через слизистые оболочки, начинается с примирования лимфоцитов, главным образом в пейеровых бляшках.
Разные лимфоидные органы защищают различные системы организма: селезенка отвечает на антигены, циркулирующие в крови; лимфоузлы реагируют на антигены, поступающие по лимфатическим сосудам; лимфоидная ткань слизистых оболочек защищает слизистые оболочки.
Лимфоциты в большинстве не оседлые, а циркулирующие клетки; они постоянно мигрируют из кровотока в лимфоидные органы и вновь поступают в кровоток.
In vivo сложные клеточные взаимодействия, составляющие основу иммунной реакции, происходят в периферических, или вторичных, лимфоидных органах, к которым относятся лимфатические узлы, селезенка и скопления диффузной лимфоидной ткани в слизистых оболочках дыхательных, пищеварительных и мочеполовых путей.
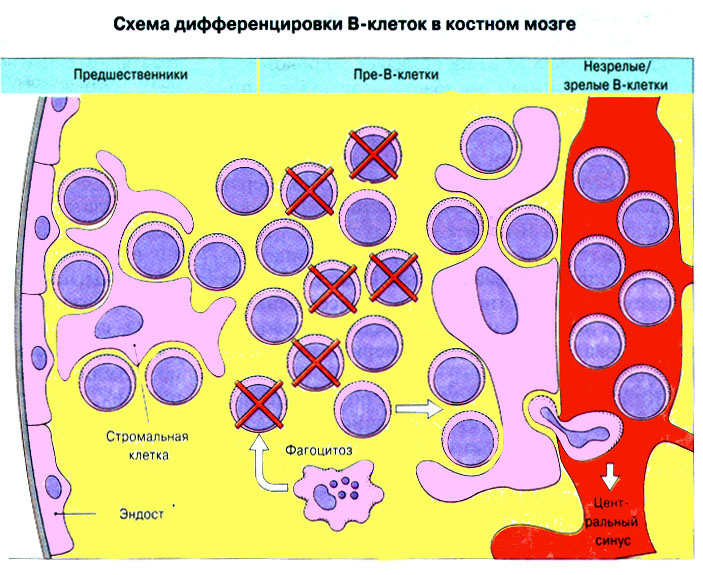
Вторичные лимфоидные ткани заселены клетками ретикулярного происхождения, а также макрофагами и лимфоцитами, предшественниками которых служат стволовые клетки костного мозга. Стволовые клетки дифференцируются в иммунокомпетентные T- и B- лимфоциты. При этом T- лимфоциты дифференцируются в иммунокомпетентные клетки в тимусе, а B- лимфоциты в костном мозге. В дальнейшем лимфоциты заселяют лимфоидные ткани, где и происходит иммунный ответ (рис. 11: "Стволовые клетки (СК) костного мозга дифференцируются в первичных лимфоидных органах в иммунокомпетентные T- и B-лимфоциты, которые затем заселяют вторичные лимфоидные органы"). (Лимфоидная ткань, ассоциированная со слизистыми оболочками, защитное действие которой основано на продукции IgA , часто обозначается сокращением
Рис. 17. Локализация участков лимфоцитопоэза в человеческом организме.
ЛИМФАТИЧЕСКИЕ УЗЛЫ (nodi lympnatici ) – бобовидной формы утолщения по ходу лимфатических сосудов, где происходит антигензависимое размножение В- и Т-лимфоцитов, приобретение ими иммунной компетенции, а также очистки лимфы от посторонних частиц. Общая масса лимфатических узлов составляет 1% массы тела, то есть около 700 г. Лимфатические узлы образуют свыше 50 групп. По топографии они подразделяются на узлы тела (соматические), внутренностей (висцеральные) и смешанные, собирающие лимфу как от внутренностей, так и других органов. Размер лимфатических узлов находится в пределах 5-10 мм.
Лимфатический узел покрыт соединительнотканной капсулой, от которой внутрь органа отходят соединительнотканные перегородки, – трабекулы. В капсуле некоторых лимфатических узлов найдены гладкие миоциты, которые принимают участие в формировании опорно-сократительного аппарата узла.
Рис. 18. Схема строения лимфатического узла. В правой части рисунка показана васкуляризация структурных элементов органа.
Паренхима узла образована В- и Т-лимфоцитами, остов для которых формирует ретикулярная ткань. Различают корковое и мозговое вещества лимфоузла. Корковое вещество образовано размещенными под капсулой лимфатическими фолликулами (узелками) – шаровидной формы скоплениями В-лимфоцитов диаметром 0,5-1 мм. Кроме В-лимфоцитов, в состав фолликулов лимфоузла входят как типичные макрофаги, так и особенная своя разновидность, которая имеет название дендритных клеток. Внешне фолликул покрыт ретикулоэндотелиоцитами – клетками, которые совмещают морфологию ретикулярных клеток с функцией эндотелия, поскольку они выстилают синусы лимфатических узлов. Среди ретикулоэндотелиоцитов есть значительное количество фиксированных макрофагов, так называемых береговых клеток. Каждый фолликул содержит светлый (реактивный, или герминативный) центр, где осуществляется размножение лимфоцитов и где локализованы преимущественно В-лимфобласты, и темную периферическую зону, в которой компактно расположены малые и средние лимфоциты. Увеличение количества и размеров реактивных центров фолликулов лимфатических узлов свидетельствует об антигенной стимуляции организма.
Рис. 19. Световая микрофотография фрагмента лимфатического узла, х 200. Окраска гематоксилин-эозином.
Мозговое вещество лимфатического узла образовано мозговыми тяжами – лентовидной формы скоплениями В-лимфоцитов, плазмоцитов и макрофагов, вытянутых в направлении от ворот узла к фолликулам. Внешне мозговые тяжи, так же, как и фолликулы коркового вещества, покрыты ретикулоэндотелиоцитами. Между мозговыми тяжами и фолликулами, соответственно, между мозговым и корковым веществами лимфатического узла размещено диффузное скопление Т-лимфоцитов, которое имеет название паракортикальной зоны. Макрофаги в составе паракортикальной зоны представлены разновидностью так называемых интердигитирующих клеток, которые контактируют между собой отростками пальцеобразной формы и производят вещества, стимулирующие пролиферацию Т-лимфоцитов. Таким образом, корковое и мозговое вещества являются бурсазависимыми, а паракортикальный слой – тимусзависимой зоной лимфатического узла.
Рис. 20. Световая микрофотография мозгового вещества лимфатического узла. Окраска гематоксилин-эозином. Светлые мозговые синусы разграничены темными мозговыми тяжами, содержащие антигенстимулированные В-лимфоциты и их эффекторные клетки - плазмоциты.
Между слоями ретикулоэндотелиоцитов, покрывающих лимфатические фолликулы и мозговые тяжи с одной стороны и соединительнотканную строму (капсулу и трабекулы) – со второго, есть щелевидные промежутки, которые называются синусами лимфатического узла. К системе синусов принадлежат краевой (размещенный между капсулой и фолликулами), вокругфолликулярные корковые синусы (между фолликулами и трабекулами), мозговые (между мозговыми тяжами и трабекулами) и воротный (в участке вгибающейся части – ворот лимфатического узла) синусы. В системе синусов осуществляется циркуляция лимфы от краевого синуса, куда впадают приносные лимфатические сосуды, через промежуточные синусы по направлению к синусу ворот, откуда лимфа будет оттекать системой выносных лимфатических сосудов. При этом лимфа очищается благодаря фагоцитозу посторонних частиц береговыми макрофагами; лимфа обогащается иммунокомпетентными Т- и В-лимфоцитами, клетками памяти, а также иммуноглобулинами (антителами).
Рис. 21. Световая микрофотография периферийного фрагмента лимфатического узла, х 400. Окраска гематоксилин-эозином. Стрелками указаны стромальные клетки - ретикулоэндотелиоциты.
Рис. 22. Световая микроскопия лимфатического узла: А – общий план строения, х 30; Б – лимфоидный фолликул со светлым реактивным центром, х 200; В – мозговой тяж в окружении ретикулярной стромы, х 200.
Механизмы функционирования лимфатического узла предусматривают тесную взаимосвязь всех его структурных компонентов. Береговые клетки и типичные макрофаги фолликулов фагоцитируют посторонние частицы, которые с лимфой проходят через систему синусов лимфатического узла. При этом при участии лизосомных ферментов макрофагов осуществляется превращение антигенов фагоцитированных частиц из корпускулярной формы в молекулярную, способную вызывать иммунный ответ: пролиферацию лимфоцитов, превращения В-лимфоцитов в плазмоциты (антителопродуценты), Т-лимфоцитов в эффекторы (Т-киллеры) и Т-клетки памяти. Активированные антигенами В-лимфоциты из фолликулов перемещаются в мозговые тяжи, превращаются там в плазмоциты – продуценты антител. Клетки памяти выходят в сосудистое русло: из них формируются эффекторные клетки после вторичного контакта с антигеном.
Рис. 23. Световая микрофотография фолликула лимфатического узла, х 400. Окраска гематоксилин-эозином. Видно крупные дендритные клетки, которые стимулируют антигензависимое размножение В-лимфоцитов.
Дендритные клетки фолликулов коркового вещества – это разновидность макрофагов, которые способны фиксировать на своей поверхности комплексы антител с антигенами. При контакте с дендритными клетками В-лимфоциты стимулируются к выработке антител. Интердигитирующие клетки паракортикальной зоны выделяют биологически активные вещества, которые стимулируют пролиферацию и созревание Т-лимфоцитов , превращения их в эффекторные клетки (Т-киллеры).
Рис. 24. Световая микрофотография фрагмента фолликула лимфатического узла, х 1000. Окраска гематоксилин-эозином. Видно крупные иммуноциты – В-лимфобласты, размножающиеся в герминативном центре под влиянием стимулирующего действия дендритных макрофагов.
Появление лимфатических узлов отмечено в конце второго месяца эмбрионального развития в виде зон локальных скоплений клеток мезенхимы вокруг лимфатических сосудов. Из внешнего слоя мезенхимы формируются капсула и трабекулы, из внутреннего – ретикулярная строма узлов. Выселения лимфобластов и лимфоцитов из костного мозга обеспечивает формирование в конце четвертого месяца эмбриогенеза мозговых тяжей и лимфатических фолликулов. Немного позже заселяется тимусзависимая паракортикальная зона и лимфатические узлы обогащаются макрофагами. В конце пятого месяца лимфатические узлы приобретают морфологические признаки, характерные для взрослого организма. Свое формирование они заканчивают на протяжении первых трех лет жизни ребенка. Реактивные центры в фолликулах появляются при иммунизации организма в процессе жизнедеятельности и становления его защитных функций. В старческом возрасте количество реактивных центров в фолликулах лимфоузлов уменьшается, падает фагоцитарная активность макрофагов, часть узлов атрофируется и происходит их замещение жировой тканью.
ГЕМОЛИМФАТИЧЕСКИЕ УЗЛЫ (nodi lymphatic haemalis ) – особенная разновидность лимфатических узлов, в синусах которых циркулирует не лимфа, а кровь, и которые выполняют функцию как лимфоидного, так и миелоидного кроветворения. У человека гемолимфатические узлы размещены в околопочечной клетчатке, вокруг брюшной аорты, реже – в заднем средостении. По строению они напоминают типичные лимфатические узлы, однако для них характерны меньшие размеры, более слабое развитие мозговых тяжей и фолликулов коркового вещества. С возрастом отмечена инволюция гемолимфатических узлов: корковое и мозговое вещества замещаются жировой клетчаткой или рыхлой волокнистой соединительной тканью.
СЕЛЕЗЕНКА (splen, lien ) – непарный орган, размещенный в брюшной полости. Селезенка имеет вытянутую форму, локализуется в левом подреберье. Масса ее 100-150 г, размеры 10х7х5 см. В селезенке осуществляется размножение и антигензависимая дифференциация лимфоцитов, а также элиминация эритроцитов и тромбоцитов, которые окончили свой жизненный цикл. Селезенка выполняет также функцию депо крови и железа, производит биологически активные вещества (спленин, фактор угнетения эритропоэза), в эмбриональном периоде является универсальным кроветворным органом. Селезенка покрыта соединительнотканной капсулой, от которой внутрь органа прорастают перегородки – трабекулы. Капсула и трабекулы, кроме богатой коллагеновыми и эластичными волокнами соединительной ткани, содержат пучки гладких миоцитов и являются опорно-сократительным аппаратом селезенки. В паренхиме селезенки различают красную и белую пульпу.
Рис. 25. Световая микрофотография селезенки крысы. Окраска гематоксилин-эозином. Количественно преобладающая красная пульпа на препарате ярко-розового цвета (оксифильная), белая пульпа – интенсивно базофильная, представляет собой скопление лимфоцитов.
Рис. 26.
Белая пульпа составляет около 20% массы органа и образована лимфоцитами, плазмоцитами, макрофагами, дендритными и интердигитирующими клетками, каркасом для которых служит ретикулярная ткань. Шарообразные скопления перечисленных видов клеток имеют название лимфатических фолликулов (узелков) селезенки. Диаметр фолликулов 0,3-0,5 мм, они окружены капсулой из ретикулоэндотелиальных клеток.
Рис. 27. Полусхематическое отображение световой микроскопии фрагмента селезенки. Окраска гематоксилин-эозином.
Лимфатический фолликул селезенки (Мальпигиево тельце) имеет четыре зоны: периартериальную, мантийную, краевую, а также светлый (реактивный, или герминативный) центр. Реактивные центры лимфатических фолликулов селезенки и лимфатического узла идентичные по структуре и функции образования. В их составе содержатся В-лимфобласты, типичные макрофаги, дендритные и ретикулярные клетки. Появление реактивных центров в фолликулах является реакцией на антигенную стимуляцию. Периартериальная зона являет собой скопление Т-лимфоцитов вокруг артерии лимфатического фолликула, или, как ее еще называют, центральной артерии селезенки. Периартериальная зона обогащена интердигитирующими клетками – макрофагами, способными фиксировать на своей поверхности комплексы антител с антигенами и вызывать пролиферацию и созревание Т-лимфоцитов. Периартериальна зона фолликулов селезенки - аналог тимусзависимой паракортикальной зоны лимфатических узлов. Темная мантийная зона образована компактно размещенными малыми В-лимфоцитами и незначительным количеством Т-лимфоцитов, плазмоцитов и макрофагов. Краевая зона – место перехода белой пульпы в красную – образована В- и Т-лимфоцитами, макрофагами и ограничена синусоидными гемокапиллярами пористого типа. После созревания лимфоцитов происходит их переход из светлого центра и периартериальной зоны в мантийную и краевую зоны со следующим выходом в кровяное русло.
Рис. 28. Световая микрофотография фрагмента Мальпигиевого тельца, х 400. Окраска гематоксилин-эозином. В центре фолликула видно светлый герминативный центр, на периферии которого находится центральная артерия.
Краевая зона, за счет контактов с гемокапиллярами, накапливает из крови большое количество антигенов и, следовательно, играет важную роль в иммунологической активности селезенки. Большое количество пульпарных артериол, отходящих от центральной артерии, покидают белую пульпу, но затем возвращаются обратно и впадают в синусы краевой зоны, окружающие узел. Большое количество макрофагов и ретикулоэндотелиальных клеток по периферии фолликула служат для фагоцитоза и удаления антигенного мусора из крови. Дендритные клетки, расположенные здесь, поглощают и передают антигены иммунологически компетентным клеткам (Т- и В-лимфоцитам), которые выходят из системного кровотока в белую пульпу именно с синусоидных капилляров краевой зоны фолликула. Активированные лимфоциты мигрируют к герминативному центру узелка, превращаются в иммунобласты (происходит т.н. бласттрансформация лимфоцитов), пролиферируют и превращаются в эффекторные клетки. Последние выходят в красную пульпу, где плазмоциты формируют скопления в виде тяжей Бильротта и продуцируют антитела, высвобождаемые в кровь. Активированные Т-лимфоциты покидают красную пульпу и возвращаются в общую циркуляцию.
Рис. 29. Тимус-зависимая и тимус-независимая зоны селезёнки. Скопление T–лимфоцитов (светлые клетки) вокруг артерий, вышедших из трабекул, образует тимус-зависимую зону. Лимфатический фолликул и окружающая его лимфоидная ткань белой пульпы - тимус-независимая зона. Здесь присутствуют B–лимфоциты (тёмные клетки), макрофаги и фолликулярные отростчатые клетки.
Лимфатические периартериальные влагалища – это удлиненной формы скопления лимфоцитов, которые в виде муфт охватывают артерии белой пульпы и с одной стороны продолжаются в лимфатические фолликулы селезенки. В центральной части влагалища, ближе к просвету сосуда, концентрируются В-лимфоциты и плазмоциты, на периферии – Т-лимфоциты.
Красная пульпа, которая составляет около 80 % массы селезенки, – это скопление форменных элементов крови, которые содержатся или в окружении ретикулярных клеток, или в системе сосудистых синусов селезенки. Участки красной пульпы, локализованные между синусами, называют пульпарными тяжами селезенки. В них осуществляются процессы превращения В-лимфоцитов в плазмоциты, а также моноцитов в макрофаги. Макрофаги селезенки способны узнавать и разрушать старые или поврежденные эритроциты и тромбоциты. При этом гемоглобин разрушенных эритроцитов утилизируется и становится источником железа для синтеза билирубина и трансферрина. Молекулы последнего изымаются из кровообращения макрофагами красного костного мозга и используются в процессе новообразования эритроцитов.
Рис. 30. Световая микрофотография фрагмента красной пульпы селезенки, х 1000. Окраска гематоксилин-эозином. Видно многочисленные макрофаги в окружении форменных элементов крови. Бледно окрашенные клетки отростчатой формы – стромальные элементы селезенки – ретикулярные клетки.
Рис. 31. Электронная микрофотография фрагмента красной пульпы селезенки. Видно пористую эндотелиальную стенку венозных синусов и лимфоидные тяжи Бильротта между ними.
Рис. 32. Сканирующая электронная микроскопия. Фрагмент венозного синуса селезенки. Видно щели между эндотелиоцитами, в которые протискиваются форменные элементы крови. Буквами обозначено: N - нейтрофил; M - макрофаг, L - лимфоцит. Макрофаги осуществляют контроль качества клеток, возвращающихся в кровеносное русло.
Сосудистая система селезенки имеет ряд особенностей, которые обеспечивают выполнение функций этого органа. Следовательно, в ворота селезенки входит селезеночная артерия, которая разветвляется на систему размещенных в трабекулах селезенки ветвей, имеющих название трабекулярных артерий. Трабекулярные артерии разделяются на артерии белой пульпы селезенки, вокруг которых группируются лимфоциты и формируются периартериальные лимфатические влагалища и фолликулы селезенки. Те части артерий белой пульпы, которые проходят через лимфатические фолликулы, имеют название центральных артерий, поскольку они служат центрами выселения лимфоцитов в процессе образования лимфатических фолликулов в онтогенезе. Центральные артерии переходят в артерии красной пульпы, последние распадаются на кисточковые артериолы, которые заканчиваются эллипсоидными (гильзовыми) артериолами. Эллипсоидные артериолы окружены своеобразными «гильзами» – муфтами из скоплений ретикулярных клеток и ретикулярных волокон, играющих роль артериальных сфинктеров селезенки. Через систему гемокапилляров эллипсоидные артериолы сообщаются с венозными синусами селезенки пористого типа. Это так называемая система закрытого (замкнутого) кровообращения селезенки. Часть капилляров, однако, может открываться непосредственно в красную пульпу, формируя систему открытого (незамкнутого) кровообращения селезенки. Венозные синусы при значительном кровенаполнении могут служить депо крови. Из венозных синусов кровь впадает в вены красной пульпы, дальше – в трабекулярные вены, а из последних – в селезеночную вену. В стенке венозных синусов в участке перехода их у вены красной пульпы имеется скопление гладких миоцитов, которые формируют венозные сфинктеры селезенки.
Рис. 33. Схема кровоснабжения селезенки.Трабекулярные артерии → пульпарные артерии → артериолы и капилляры фолликула → синусы краевой зоны → выход Т- и B–лимфоцитов из сосудистого русла. Артериолы фолликула → кисточковые артериолы красной пульпы → капилляры-синусоиды.
Рис. 34. Синусоиды в красной пульпе селезёнки. По теории незамкнутой циркуляции (вверху), кровь из капилляров поступает в красную пульпу, а затем - в синусоиды. По теории замкнутой циркуляции (снизу), капилляры открываются прямо в синусоиды.
Рис. 35. Световая микроскопия фрагментов селезенки с использованием различных гистологических красителей: А – импрегнация азотистокислым серебром (видно ретикулярную строму); Б – окраска гематоксилин-эозином (визуализируются структурные компоненты паренхимы органа); В – окраска железным гематоксилином (четко видна разная плотность лимфоидных элементов в белой пульпе).
При сокращении венозных сфинктеров, кровь скапливается в синусах, происходит ее згущение в результате пропитки плазмы через стенку венозных синусов. При одновременном сокращении артериальных и венозных сфинктеров наблюдается депонирование крови в селезенке. Расслабление артериальных и венозных сфинктеров при одновременном сокращении гладких миоцитов капсулы и трабекул селезенки предопределяет выброс депонируемой крови в венозное русло.
Закладка селезенки осуществляется в начале второго месяца эмбрионального развития в виде пронизанных сосудами скоплений клеток мезенхимы в дорсальной брыжейке. Из мезенхимы формируется ретикулярная ткань, последнюю заселяют стволовые клетки крови. На третьем месяце эмбриогенеза в селезенке дифференцируется периартериальная тимусзависимая зона, на пятом месяце формируются реактивные центры и краевые зоны фолликулов, на шестом месяце можно различить красную пульпу. В это же время (с третьего до пятого месяцев эмбриогенеза) в селезенке нарастают явления миелоидного гемопоэза, она выполняет функции универсального кроветворного органа. Начиная с шестого месяца и к рождению ребенка проявления миелоидного кроветворения угасают, их вытесняют процессы лимфоцитопоэза. В зрелом возрасте селезенка проявляет значительные репаративные возможности; экспериментально доказана возможность ее возобновления при потере 80–90% паренхимы. Масса селезенки несколько уменьшается в возрасте с 20 до 30 лет; в промежутке с 30 до 60 лет она стабильна. В старческом возрасте отмечена атрофия красной и белой пульп, разрастания соединительнотканной стромы, снижение содержания среди паренхиматозных элементов макрофагов и лимфоцитов, повышение содержания гранулоцитов и тканевых базофилов, появление мегакариоцитов. Ухудшается утилизация железа из разрушенных в селезенке эритроцитов.
Межклеточные взаимодействия
в обеспечении иммунной защиты организма.
Для адекватной реакции на посторонние вещества, которые попадают в организм (антигенную стимуляцию), необходимо взаимодействие и кооперация разных видов клеток иммунной системы. Среди них различают клетки макрофагической природы – моноциты крови, гистиоциты -макрофаги соединительной ткани, костномозговые, перитонеальные, альвеолярные макрофаги , клетки Лангерганса кожи, клетки Кащенко-Хофбауэра плаценты, звездчатые ретикулоэндотелиоциты печени, дендритные и интердигитирующие клетки лимфатических узлов и селезенки, остеокласты костной ткани, микроглиоциты нервной системы. Есть группа так называемых микрофагов, к которым принадлежат нейтрофильные гранулоциты крови, а также клетки, которые при определенных условиях функционирования могут проявлять фагоцитарные свойства, – эндотелиоциты , фиброкласты . Наконец, третья группа клеток объединяет разные популяции Т- и В-лимфоцитив (Т-киллеры, Т-хелперы, Т-супрессоры, плазмоциты, Т- и В-клетки памяти ). Общая масса клеток, которые непосредственно обеспечивают иммунную защиту организма, составляет около 1% массы тела.
На проникновение посторонних частиц в организм прежде всего реагируют Т-хелперы: происходит связывание антигенных детерминант со специфическими рецепторами на их поверхности. Образованный антигенрецепторный комплекс отрывается от поверхности плазмолемы Т-хелпера и фиксируется поверхностными рецепторами макрофага. На следующем этапе модифицированные макрофагами антигены передаются В-лимфоцитам, которые под воздействием антигенной стимуляции и активирующего действия Т-хелперов превращаются в плазмоциты. Последние синтезируют белковые молекулы иммуноглобулинов (антител), которые избирательно связываются с антигенами и предопределяют их инактивацию. Т-хелперы после контакта с антигеном производят особенные химические вещества, которые стимулируют пролиферацию Т-киллеров. Последние имеют способность разрушать клеточные оболочки бактерий и клеток, которые несут на своей поверхности антигенные детерминанты.
На каждом из перечисленных этапов может происходить частичная инактивация постороннего материала, а также его модификация и передача другим популяциям клеток для выработки иммунного ответа. Возможным является вариант, когда антигенсодержащая частица распознается и увлекается макрофагом без участия Т-лимфоцита, расщепляется его лизосомными ферментами, а полученные антигенные фрагменты передаются Т- и В-лимфоцитам и стимулируют их превращение в эфекторные клетки (Т-киллеры и плазмоциты), а также клетки памяти.
Взаимодействие клеток при иммунном ответе
Иммунный ответ возможен в результате активации клонов лимфоцитов и состоит из двух фаз. В первой фазе антиген активирует те лимфоциты, которые его распознают. Во второй (эффекторной) фазе эти лимфоциты координируют иммунный ответ, направленный на устранение антигена.
Гуморальный иммунный ответ
В гуморальном иммунном ответе участвуют макрофаги (антигенпредставляющие клетки), T–хелперы и B–лимфоциты.
Макрофаг поглощает вторгшийся в организм антиген и подвергает его процессингу – расщеплению на фрагменты. Фрагменты антигена выставляются на поверхности клетки вместе с молекулой MHC. Комплекс «антиген–молекула MHC класса II» предъявляется T–хелперу (рис. 35).
Рис. 36. Распознавание антигена рецептором T–лимфоцита . При помощи рецептора T–лимфоцита T–клетка распознает антиген, но только находящийся в комплексе с молекулой MHC. В случае Т-хелпера в процессе участвует её молекула – CD4, которая свободным концом связывается с молекулой MHC. Распознаваемый T–клеткой антиген имеет два участка: один взаимодействует с молекулой MHC, другой (эпитоп) связывается с рецептором T–лимфоцита. Подобный тип взаимодействия, но с участием молекулы CD8, характерен для процесса распознавания T-киллером антигена, связанного с молекулой MHC класса I.
T–хелпер распознаёт комплекс «антиген–молекула MHC класса II» на поверхности антигенпредставляющей клетки. Процесс распознавания включает взаимодействие комплекса T–клеточный рецептор–СD3, обеспечивающего специфичность и участие вспомогательных костимулирующих молекул. Для активации T–хелпера специфическое узнавание T–хелпером фрагмента антигена на поверхности антигенпредставляющей клетки оказывается недостаточным. Активацию T–хелперов обеспечивает взаимодействие молекулы В7 (СD80) на поверхности антигенпредставляющей клетки с молекулой CD28 на поверхности T–хелпера. К cтимуляции через CD28 особенно чувствительны T–хелперы, которые дифференцируются в T–хелперы 2, активирующие B–клетки через СD80. При слабой экспрессии СD28 и в присутствии молекулы CTLA (C ytotoxic T - l ymphocyte p rotein ) формируются T–хелперы 1.
Узнавание T–хелпером нужных молекул на поверхности антигенпредставляющей клетки стимулирует секрецию ИЛ1. Активированный ИЛ1 T–хелпер синтезирует ИЛ2 и рецепторы ИЛ2, через которые агонист стимулирует пролиферацию T–хелперов и цитотоксических T–лимфоцитов. В случае T–хелпера речь идёт об аутокринной стимуляции, когда клетка реагирует на тот агент, который сама же синтезирует и секретирует. Таким образом, после взаимодействия с антигенпредставляющей клеткой T–хелпер приобретает способность отвечать на действие ИЛ2 всплеском пролиферации. Биологический смысл этого процесса состоит в накоплении такого количества T–хелперов, которое обеспечит образование в лимфоидных органах необходимого количества плазматических клеток, способных вырабатывать антитела против данного антигена.
B–лимфоцит . Активация B–лимфоцита предполагает прямое взаимодействие антигена с иммуноглобулином (Ig) на поверхности B–клетки. В этом случае сам B–лимфоцит процессирует антиген и представляет его фрагмент в связи с молекулой MHC II на своей поверхности. Этот комплекс распознаёт T–хелпер, отобранный при помощи того же антигена, который участвовал в отборе данного B–лимфоцита. В активации B–клетки участвуют две пары молекул: с одной стороны специфическое взаимодействие антигена с рецептором Ig M на поверхности B–лимфоцита, а с другой стороны молекула CD40 на поверхности B–клетки взаимодействует с молекулой CD40L (CD154) на поверхности T–хелпера, активирующего B–клетку. Узнавание рецептором T–хелпера комплекса «антиген–молекула MHC класса II» на поверхности B–лимфоцита приводит к секреции из T–хелпера ИЛ2, ИЛ4, ИЛ5 и γ-ИФН, под действием которых B–клетка активируется и пролиферирует, образуя клон. Активированный B–лимфоцит дифференцируется в плазматическую клетку: увеличивается количество рибосом, гранулярная эндоплазматическая сеть и комплекс Гольджи становятся более выраженными.
Плазматическая клетка синтезирует Ig. ИЛ6, выделяемый активированными T–хелперами, стимулирует секрецию Ig. Часть зрелых B–лимфоцитов после антиген-зависимой дифференцировки циркулирует в организме как клетки памяти.
Клеточный иммунный ответ
Клеточный иммунный ответ характеризуется пролиферацией коммитированных иммунокомпетентных клеток, реагирующих с антигеном в комплексе с молекулой MHC класса I на поверхности чужеродных клеток или эндогенными антигенами в комплексе с молекулой MHC класса I на поверхности собственных вирус-инфицированных и трансформированных (опухолевых) клеток. В клеточном иммунном ответе участвует цитотоксический T–лимфоцит.
Реакции клеточно-опосредованного цитолиза . Эффекторные клетки при помощи своих рецепторов узнают клетку-мишень и уничтожают её. За клеточно-опосредованный цитолиз отвечают не только T–лимфоциты, но и другие субпопуляции лимфоидных клеток и в некоторых случаях миелоидные клетки. В процессе узнавания участвуют различные молекулы, выставленные на поверхности взаимодействующих клеточных партнёров:
√ специфические антигены, например, вирусные пептиды на поверхности инфицированных клеток, в комплексе с молекулой MHC распознаются рецепторами цитотоксических T–клеток, преимущественно CD8 + - и некоторыми субпопуляциями CD4 + -клеток;
√ антигенные детерминанты опухолевых клеток распознаются NK–клетками без участия молекулы MHC класса I;
√ связанные с антигеном АТ на поверхности клеток-мишеней, распознаются рецепторами Fc–фрагментов NK–клеток (феномен АТ-зависимой цитотоксичности).
Цитотоксический T–лимфоцит (T C). Предъявленный на поверхности клетки-мишени антиген в комплексе с молекулой MHC класса I связывается с рецептором цитотоксического T–лимфоцита. В этом процессе участвует молекула CD8 клеточной мембраны T C . Секретируемый T–хелперами ИЛ2 стимулирует пролиферацию цитотоксических T–лимфоцитов.
Уничтожение клетки -мишени . Цитотоксический T–лимфоцит раcпознаёт клетку-мишень и прикрепляетcя к ней. В цитоплазме активированного цитотоксического T–лимфоцита присутствуют мелкие гранулы. В них содержится цитолитический белок перфорин. Выделяемые T-киллером молекулы перфорина полимеризуютcя в мембране клетки-мишени в приcутcтвии Ca 2+ . Сформированные в плазматической мембране клетки-мишени перфориновые поры пропуcкают воду и cоли, но не молекулы белка. Если полимеризация перфорина произойдет во внеклеточном проcтранcтве или в крови, где в избытке имеетcя кальций, то полимер не cможет проникнуть в мембрану и убить клетку. Сам T-киллер защищён от цитотокcичеcкого дейcтвия перфорина.
На практической работе предлагаются к изучению следующие гистологические препараты:
1. МАЗОК КРАСНОГО КОСТНОГО МОЗГА.
Окраска: согласно Романовского- Гимзы (азур II, еозин).
При малом, а затем при большом увеличении микроскопа найти в препарате синусоидные капилляры. В их просвете видно эритроциты и лейкоциты. Между синусоидными капиллярами находится ретикулярная ткань, в петлях которой размещаются гемопоэтические клетки на разных стадиях созревания. Клетки эритропоэтического ряда: а) проэритробласты – крупные клетки (15 мкм) с большим округлым ядром, в котором хорошо видно ядрышки. Сильно базофильная цитоплазма узкой полоской окружает ядро; б) базофильные эритробласты – клетки (10-12 мкм) с базофильной цитоплазмой. Ядро округлое, содержит больше гетерохроматина и имеет темную окраску; в) полихроматофильные эритробласты – еще более мелкие клетки (8-10 мкм), их цитоплазма окрашивается одновременно как кислыми, так и щелочными, красителями и выглядит серовато-зеленоватой. Ядра интенсивно окрашены и не имеют ядрышек; г) оксифильные эритробласты (нормобласты) – цитоплазма клеток содержит уже значительное количество гемоглобина, поэтому окрашивается оксифильно, ядра пикнотизируются, часто лизируются и исчезают; д) ретикулоциты – молодые эритроциты. В цитоплазме этих клеток можно видеть зернисто-сетчатые структуры – остатки ядра и органел; ж) зрелые эритроциты – безъядерные форменные элементы крови, с гомогенной оксифильной цитоплазмой.
Клетки гранулоцитопоэтического ряда: а) промиелоциты – крупные клетки с большими округлыми и светлыми ядрами. Цитоплазма умеренно базофильна, в ней содержатся одиночные азурофильные гранулы (лизосомы); б) миелоциты содержат в цитоплазме специфическую зернистость: эозинофильные имеют крупные ярко красные эозинофильные гранулы; базофильные – темно-синие базофильные гранулы; нейтрофильные – имеют два вида мелких зерен, которые воспринимают как основные, так и кислые красители. Миелоциты в процессе созревания превращаются в метамиелоциты. Характерным морфологическим признаком этой переходной формы клеток является: уменьшение объема цитоплазмы и ядра, которые приобретают вид выгнутой палочки или подковы. В зависимости от гранул, содержащихся в цитоплазме, метамиелоциты также подразделяются на базофильные, оксифильные и нейтрофильные. По ходу созревания ядра метамиелоцитов сегментируются и клетки превращаются в зрелые сегментоядерные эозинофилы, базофилы и нейтрофилы.
В мазке красного костного мозга около синусоидов локализуются мегакариоциты – очень большие (> 50 мкм) клетки с несколькими полиплоидными ядрами и базофильной цитоплазмой.
Изучить препарат, зарисовать и обозначить: 1. Проэритробласты. 2. Полихроматофильные эритробласты. 3. Нейтрофильные метамиелоциты. 4. Мегакариоциты. 5. Зрелые клетки крови (эритроциты, лейкоциты) в синусоидном гемокапилляре.
По каким морфологическим признакам можно отличить полихроматофильный проэритроцит от эритроцита?
На каком этапе дифференциации миелоцита можно определить базофильные, оксифильные и нейтрофильные клетки?
В мазке красного костного мозга какая клетка имеет больший размер: мегакариобласт или мегакариоцит?
На основании изучения мазка красного костного мозга назовите клетки, которые по размерам отвечают эритроцитам.
На каком этапе дифференциации миелоцит теряет способность к делению?
2. ЗОБНАЯ ЖЕЛЕЗА (Тимус).
При малом увеличении видно дольковое строение железы. В каждой дольке есть более темная периферийная часть – корковое вещество и более светлая внутренняя – мозговое вещество. При большом увеличении в средней части мозгового вещества видно эпителиальные тельца Гассаля – концентрическое наслоение эпителиоретикулоцитов. Стромальными элементами дольки являются эпителиоретикулоциты, которые контактируют своими отростками, образуя сеть. В промежутках между эпителиоретикулоцитами расположены паренхиматозные элементы: Т-лимфоциты (тимоциты) и макрофаги.
Изучить препарат, зарисовать и обозначить: 1. Долька железы: 1. Корковое вещество: а) Т-лимфоциты; 2. Мозговое вещество: б) эпителиоретикулоциты; в) тельца Гассаля. ІІ. Междольковая соединительная ткань. 3. Кровеносные сосуды.
На основании изучения препарата тимуса сделайте вывод об интенсивности митозов в субкапсулярном участке, корковом и мозговом веществе.
Что такое тельца Гассаля и где они локализуются в тимусе?
Эпителий, который образует строму тимуса однослойный или многослойный? Объясните почему.
3. ЛИМФАТИЧЕСКИЙ УЗЕЛ.
При малом увеличении микроскопа видно, что лимфатический узел покрыт соединительнотканной капсулой, от которой внутрь отходят тонкие перегородки, трабекулы. Между трабекулами размещается ретикулярная ткань, инфильтрованная многочисленными лимфоцитами. Лимфоциты сосредотачиваются по периферии узла в виде больших скоплений округлой формы – фолликулов, которые образуют корковое вещество лимфатического узла. От фолликулов в глубину узла отходят мозговые тяжи, которые формируют мозговое вещество. Светлые промежутки, заполненные ретикулярной тканью и небольшим количеством лимфоцитов, являются синусами. Краевой синус, располагающийся между фолликулами и капсулой, переходит в промежуточные корковые синусы, а те в свою очередь продолжаются в промежуточные мозговые, которые собирают лимфу в центральный синус у ворот лимфоузла.
Зарисовать препарат и обозначить: 1.Капсула. 2.Трабекулы. 3. Корковое вещество. 4. Мозговое вещество. 5. Фолликулы. 6.Мозговые тяжи. 7.Краєвой синус. 8. Промежуточные корковые синусы. 9. Промежуточные мозговые синусы. 10. Ворота лимфатического узла. 11. Ретикулярная ткань.
На основании изученного препарата сделайте вывод об антигенной стимуляции лимфатического узла и объясните свой ответ.
Что циркулирует в синусах лимфатического узла?
Укажите место на препарате, где больше всего находится плазмоцитов.
4. СЕЛЕЗЕНКА.
Окраска: гематоксилином эозином.
При малом увеличении микроскопа хорошо видно плотную соединительнотканную капсулу, от которой внутрь органа отходят трабекулы. Капсула селезенки покрыта мезотелием и содержит небольшое количество гладкомышечных клеток, которые сокращаясь способствуют выбросу крови через ворота органа. Между трабекулами размещается белая и красная пульпа селезенки. Белая пульпа состоит из лимфатических фолликулов, которые имеют центральную артерию. Красная пульпа образована ретикулярной тканью, форменными элементами крови, преимущественно эритроцитами и венозными синусами.
Изучить препарат при большом увеличении, зарисовать небольшой участок и обозначить: 1. Капсула селезенки. 2.Трабекулы. 3. Белая пульпа (лимфатические фолликулы): а) центральная артерия; б)герминативный центр. 4.Красная пульпа: а) ретикулярная ткань, б) эритроциты; в) лейкоциты. 5.Венозный синус.
Назовите основные отличительные особенности строения лимфатических фолликулов селезенки и лимфатических узлов.
Сделайте вывод, где больше лимфоидных узелков с центрами размножения в селезенке или лимфатическом узле и объясните почему?
ДЕМОНСТРАЦИОННЫЕ ПРЕПАРАТЫ:
1. Мазок красного костного мозга ребенка 5 лет.
2. Зобная железа ребенка 12 лет.
3. Ретикулярные волокна в лимфатическом узле. Импрегнация серебром.
ЭЛЕКТРОННЫЕ МИКРОФОТОГРАФИИ:
1. Клетки красного костного мозга.
2. Эпителиоретикулоциты зобной железы.
3. Т-лимфоциты.
4. Синус лимфатического узла.
5. Синус селезенки.
Ситуативные задачи:
1. В препарате есть срез трубчатой кости ребенка 3-5 лет, юноши 12-18 лет и старого человека. Как с возрастом изменяется состояние и топография красного костного мозга?
2. У новорожденного ребенка удалили тимус. В результате этой операции у нее резко снизилась способность к продукции антител. Объяснить причину этого явления.
3. Есть микрофотографии лимфатических узлов брюшины, сфотографированные на высоте пищеварения и в состоянии покоя. Как можно отличить лимфатический узел во время пищеварения и объяснить это явление?
4. Животного сразу же после рождения поместили в стерильные условия. Могут ли в данной ситуации формироваться вторичные фолликулы в лимфатических узлах, если так, то почему, если нет, то почему?
5. В древние времена марафонцам удаляли селезенку. Объясните почему?
ЛИМФОИДНАЯ ТКАНЬ (анат. lympha, от лат. lympha чистая вода, влага + греч. -eidēs подобный) - представляет собой совокупность лимфоцитов и макрофагов, которые располагаются в клеточно-волокнистой ретикулярной строме. Лимфоидная ткань образует действующую паренхиму лимфоидных органов. Лимфоидные органы являются органами иммуногенеза, которые включают в свой состав вилочковую железу, лимфатические узлы, селезенку, лимфоидные элементы костного мозга и скопления лимфоидной ткани в стенках дыхательных, мочевыводящих путей и желудочно-кишечного тракта.
Основой лимфоидной ткани являются ретикулярные волокна и клетки, которые образуют сеть с ячейками различных размеров. Эта сеть оснащена петлями, в которых расположены клетки лимфоидного ряда, макрофаги, тучные клетки, а также незначительное количество лейкоцитов. Образование ретикулярной стромы происходит из мезенхимы, а образование клеток лимфоидного ряда из стволовых клеток костного мозга.
Клетки лимфоидного ряда подразделяются на две группы :
- Т-лимфоциты
- В-лимфоциты.
Перемещение клеток происходит с кровью и лимфой. Функция клеток лимфоидного ряда заключается в участии в реакциях иммунного ответа направленного против генетически инородных веществ.
Строение лимфоидной ткани, топография элементов ее структуры в различных органах иммунной системы отличается своими особенностями. Лимфоидная ткань в центральных органах иммуногенеза поддерживает функциональное единство с другими тканями. Например, в вилочковой железе лимфоидная ткань находится в функциональном единстве с эпителиальной тканью, а в костном мозге - с миелоидной тканью. Нахождение лимфоидной ткани в различных качественных состояниях напрямую зависит от зрелости и функционального состояния периферических органов иммунной системы, так как наличие единичных лимфоцитов и лимфоидных узелков свидетельствует о повышенной иммунной активности организма.
Самое большое количество лимфоидных узелков можно обнаружить в таких органах, как: миндалины, лимфоидные бляшки, селезенка, стенки червеобразного отростка, желудок, тонкая и толстая кишки, лимфатические узлы у детей малого и подросткового возраста.
Лимфоидная ткань образует редкий и тонкий защитный слой клеток лимфоидного ряда и размещается под эпителиальным покровом мочевыводящих и дыхательных путей, желудочно-кишечного тракта. Лимфоидная ткань способствует образованию в селезенке лимфоидных муфт вокруг артериальных сосудов. Уменьшение количества лимфоидной ткани и лимфоидных узелков зависит от процесса старения организма. Если в организме наблюдаются воспалительные процессы и активации иммунных реакций первичного и вторичного характера, следовательно, происходит реактивная гиперплазия лимфатических узлов. Поражение лимфоидной ткани происходит при гистиоцитах, парапротеинемических гемобластозах, лимфогранулематозе, гистиоцитах Х, злокачественных лимфомах.
Жизненный цикл иммунокомпетентных клеток лимфоидной ткани (В- и Т-клетки и их потомки в виде клеток, синтезирующих иммуноглобулины и антигенраспознающие и эффекторные лимфоциты) существенно короче, чем время жизни организма. В связи с этим ведущим условием для осуществления иммунологических реакций служит постоянное протекание в лимфоидной ткани гистогенезов, в ходе которых происходят пролиферация предшественников иммунокомпетеитных клеток и их дифференцировка в зрелые клетки.
Последние, как правило, высокоспециализированы и гетерогенны , а дифференцировочные различия, связанные с иммунологическими функциями, у них жестко закреплены (класс и специфичность синтезируемого иммуноглобулина у В-клеток и специфичность рецепторов для антигенов у Т-клеток). Наоборот, среди клеток-предшественников в течение всей жизни присутствуют и такие, для которых открыты несколько возможностей дифференцировки.
Орган иммунитета - - имеет еще одну важную особенность. Большинство его клеток-предшественников относится к подвижным, рециркулирующим элементам. Они образуются в местах, отдаленных от тех, куда они затем мигрируют и где образуют потомство зрелых клеток.
Таким образом, процессы , которые во многих других тканях осуществляются лишь в момент их закладки при морфогенезе, в лимфоидной ткани протекают всю жизнь. Они обеспечивают постоянное изменение ее клеточного состава, набора синтезируемых иммуноглобулинов (антител) и типов иммунологически активных лимфоцитов в соответствии с меняющимся спектром антигенов, поступающих в лимфоидную ткань.
На лимфоидной ткани удалось убедительно показать, что эти изменения осуществляются главным образом или даже исключительно путем селекции коммитированных клеток-предшественников, предетерминированных к последующей дифференцировке безотносительно к воздействию антигенов. Таким образом, в дифференцировке лимфоидных клеток во взрослом организме естественно выделяются два этапа: развитие антигеннезависимых предшественников и антигензависимая дифференцировка их потомков.
Первый этап начинается с общей стволовой кроветворной клетки и оканчивается коммитированными предшественниками. Неизвестно, однако, в какой последовательности происходят при этом стадии разделения на Т- и В-предшественники, на предшественники, различные по иммунологической специфичности и по классам синтезируемых иммуноглобулинов, а также по принадлежности к разным субпопуляциям Т-клеток. Результаты, полученные при использовании трансплантации костномозговых клеток, маркированных пострадиационными хромосомными перестройками, показали, в частности, что в костном мозге содержатся стволовые клетки, способные к самоподдержанию, но имеющие ограниченные дифференцировочные потенции,- например, такие, которые дифференцируются только в миелоидные или только в Т-клетки.
При этом не обнаружилось пока стволовых клеток , лишенных потенций к миелоидной дифференцировке, но сохранивших свойство быть предшественниками одновременно и для Т- и для В-клеток. Можно поэтому предполагать, что самоподдерживающиеся предшественники Т-клеток (претимические стволовые клетки) прямо происходят из полипотентных стволовых кроветворных клеток, не проходя стадии предшественника, общего для всех лимфоидных клеток. Что касается В-клеток, то между ними и стволовыми кроветворными клетками, возможно, нет вообще предшественников, способных к самоподдержанию.
Вопрос о механизмах, действующих при образовании коммитированных предшественников и регулирующих экспрессию в них генов, ответственных за иммунологические функции, остается открытым. Есть веские основания считать, что этот важный этап развития иммунокомпетентных клеток является антигеннезависимым. Наоборот, последующее развитие уже коммитированных клеток и их потомков отчетливо антигензависимо. Оно обеспечивает изменение клеточного состава лимфоидной ткани путем преимущественного размножения и дифференцировки тех предшественников, чьи поверхностные рецепторы распознают присутствующие в ней в данный момент антигены.
Другой механизм избирательной пролиферации коммитированных предшественников связан с клеточными взаимодействиями, при которых активированная антигеном клетка стимулирует соседние с ней клетки-предшественники. И в том и в другом случае антигены выступают как инициаторы изменения состава клеточных популяций населяющих лимфоидную ткань, и как индукторы синтезируемых этой тканью белков.
Лимфоидная ткань очень чувствительна к внешним и внутренним воздействиям. По мере старения организма уменьшается количество Л.т. и лимфоидных узелков в органах иммунной системы.
Лимфоидная ткань (синоним лимфатическая ткань) - собирательный термин для обозначения структур, в которых происходит образование лимфоцитов. Лимфоидная ткань человека составляет около 1% веса тела и является одним из важнейших компонентов лимфоидных органов.
Что такое Гипертрофия лимфоидной ткани глотки -
Одной из основных функций лимфоидных органов является их участие в процессах кроветворения (лимфопоэз). С этой способностью лимфоцитов связана важная функция лимфоидной ткани - участие ее в защитных реакциях организма. Большое влияние на степень развития лимфоидной ткани имеют гормоны коры надпочечников. Недостаточная функция коры надпочечников вызывает разрастание лимфоидной ткани. Введение гормонов коры надпочечников приводит к дегенерации лимфоидной ткани и гибели лимфоцитов.
Строение и роль лимфоидной ткани в деятельности иммунной системы
Строение Л.т., топография ее структурных элементов в различных органах иммунной системы имеет свои особенности. В центральных органах иммуногенеза Л.т. находится в функциональном единстве с другими тканями, например в костном мозге - с миелоидной тканью, в вилочковой железе - с эпителиальной тканью. Помимо скоплений, Л. т. в виде редкого, тонкого, как бы защитного слоя клеток лимфоидного ряда располагается под эпителиальным покровом дыхательных и мочевыводящих путей, желудочно-кишечного тракта.
Лимфоидная ткань слизистых оболочек: введение
Лимфоидные органы относят либо к первичным (центральным), либо ко вторичным органам. Таким образом, лимфоциты относятся к той категории клеток, которые широко распространены в организме. Лимфоидная ткань представляет собой разновидность соединительной ткани, которая характеризуется высоким содержанием лимфоцитов.
В большинстве лимфоидных органов фибробластоподобные ретикулярные клетки образуют эти волокна, на которых располагаются их многочисленные отростки. Узелковая лимфоидная ткань образована сферическими скоплениями лимфоцитов; это - так называемые лимфоидные узелки, или лимфоидные фолликулы, содержащие преимущественно В-лимфоциты. Лимфоидная ткань, ассоциированная со слизистыми оболочками, защитное действие которой основано на продукции IgA , часто обозначается сокращением MALT (mucosal-associated lymphoid tissue).
Язычная миндалина состоит из скоплений лимфоидной ткани - лимфоидных узел- ков, число которых (80-90) наиболее велико в детском, подростковом и юношеском возрасте. К моменту рождения количество лимфоидных узелков в формирующейся миндалине заметно возрастает. Центры размножения в лимфоидных узелках появляются уже вскоре после рождения (на 1-м месяце жизни) . В дальнейшем их количество увеличивается вплоть до юношеского возраста.
Строение лимфоидной ткани. Гистология, функции
К язычной миндалине подходят ветви правой и левой язычных артерий, а также, в редких случаях, ветви лицевой артерии. От этой пластинки в медиальном направлении в лимфоидную ткань органа отходят трабекулы (перегородки) , которые при хорошей их выраженности разделяют миндалину на дольки.
5-месячного плода миндалина представлена скоплением лимфоидной ткани размером до 2-3 мм. В этот период в образующуюся миндалину начинают врастать эпителиальные тяжи - формируются будущие крипты. На поверхности складок у детей видны многочисленные мелкие бугорки, в глубине которых находятся скопления лимфоидной ткани - лимфоидные узелки.
Под эпителиальным покровом в диффузной лимфоидной ткани находятся лимфоидные узелки глоточной миндалины диаметром до 0,8 мм, большинство из которых имеют центры размножения. Глоточная миндалина закладывается на 3-4-м месяце внутриутробной жизни в толще формирующейся слизистой оболочки носовой части глотки.
К концу года ее длина достигает 12 мм, а ширина - 6-10 мм. лимфоидные узелки в миндалине появляются на 1-м году жизни. После 30 лет величина глоточной миндалины постепенно уменьшается. Возрастная инволюция трубной миндалины начинается в подростковом и юношеском возрасте. Наблюдается обычно у детей в возрасте 3-10 лет. Гипертрофированная лимфоидная ткань подвергается физиологической инволюции и уменьшается в Период полового созревания.
Сохраняя свою функцию, гипертрофированная лимфоидная ткань может, однако, быть причиной патологических изменений в носу, ушах и гортани. Гипертрофия небных миндалин часто сочетается с гипертрофией всего глоточного лимфоидного кольца, особенно с гипертрофией глоточной миндалины. В период полового созревания аденоиды подвергаются обратному развитию, но возникшие осложнения остаются и часто приводят к инвалидизации. Косвенными признаками аденоидов является также гипертрофия небных миндалин и лимфоидных элементов на задней стенке глотки.
Гипертрофия лимфоидной ткани в ответ на инфекционное заболевание приводит к учащению воспалительных процессов в глотке. В толще миндалины располагаются округлые плотные скопления лимфоидной ткани - лимфоидные узелки миндалины. Участки лимфоидной ткани находятся в слизистой оболочке некоторых органов (бронхов, мочевых путей, почек).
План лекции:
Классификация, функции и общий принцип строения органов кроветворения
Понятие о лимфоидной и миелоидной ткани, развитие миелоидного кроветворения
Красный костный мозг (ККМ):
3.1. Функции ККМ
3.2. Строение ККМ
3.3. Особенности кровоснабжения ККМ
3.4. Регенерация ККМ
Тимус:
4.1. Функции тимуса
4.2. Развитие тимуса
4.3. Строение тимуса
4.4. Особенности кровоснабжения тимуса. Гемато-тимусный барьер.
4.5. Возрастные особенности тимуса
Функции органов кроветворения
Органы кроветворения и иммунной защиты образуют единую с кровью и лимфой систему, которая:
Обеспечивает непрерывный процесс обновления форменных элементов крови в результате постоянной пролиферации и дифференцировки клеток в соответствии с потребностями организма.
Создает и осуществляет комплекс защитных реакций от повреждающего действия факторов внешней и внутренней среды, иммунный надзор за деятельностью клеток своего организма.
Поддерживает целостность и индивидуальность организма благодаря способности клеток иммунной системы отличать структурные компоненты своего организма от чужеродного и уничтожать последние.
К органам кроветворения и иммуногенеза относятся:
Лимфатические и гемолимфатические узлы,
Селезенка,
Лимфоидые образования пищеварительного тракта, к которым относятся миндалины, пейеровы бляшки, аппендикс, лимфоидные образования половой, дыхательной, выделительной систем.
Красный костный мозг (ККМ),
Все органы кроветворения и иммуногенеза подразделяются на центральные и периферические .
К центральным относится ККМ и тимус. В них локализованы стволовые кроветворные клетки и происходит первый этап дифференцировки лимфоцитов, называемый антигеннезависимым .
К периферическим органам относятся: селезенка, лимфатические и гемолимфатические узлы, лимфоидные образования по ходу пищеварительной трубки, половой, дыхательной, выделительной систем. В этих органах осуществляется антигензависимая дифференцировка лимфоцитов.
Общий принцип строения органов кроветворения
Основу всех органов кроветворения формирует стромальный компонент, представленный ретикулярной тканью, исключением является лишь тимус, его стромальный компонент представлен эпителиоретикулярной тканью, имеющей эпителиальное происхождение. Клетки стромывыполняют опорную, трофическую и регуляторную функции, обладают в каждом органе характерными признаками. Они создают особое микроокружение, синтезируя гемопоэтины для правильного развития кроветворных клеток, ГАГ кислые и нейтральные, а так же белок ламинин, создающий трехмерную сеть для миграции клеток крови.
Все органы гемопоэза и иммуногенеза среди клеток стромы содержат большое количество макрофагов, которые участвуют в созревании и дифференцировке формирующихся форменных элементов, а также в фагоцитозе разрушенных клеток, учавствуя в их утилизации.
В строме органов кроветворения содержится сосудистый компонент , который представлен особыми кровеносными сосудами, синусными капиллярами, с высоким эндотелием , который, в свою очередь, обеспечивает распознавание зрелых клеток, способен сортировать их и обеспечивать миграцию форменных элементов в кровеносное русло.
В сети стромосоздающей ткани находятся форменные элементы крови на разных этапах созревания – гемопоэтический компонент .
Понятие о лимфоидной и миелоидной ткани, развитие органов миелоидного кроветворения
Кроветворные клетки в совокупности со стромой образуют два типа тканей миелоидную и лимфоидную:
Миелоидная ткань – это ретикулярная ткань, с находящимися там развивающимися клетками миелоидного ряда (эритропоэза, тромбоцитопоэза, гранулоцитопоэза, моноцитопоэза) и лимфоидного (В-лимфоцитопоэз). Миелоидная ткань формирует основу органов миелоидного кроветворения, к которым у человека относится красный костный мозг.
Лимфоидная ткань - это ретикулярная или эпителиоретикулярная ткань (тимус), в которой находятся клетки лимфоидного ряда (лимфоцитопоэза) на разных стадиях развития. Лимфоидная ткань формирует органы лимфоидного кроветворения, к которым относятся: тимус, селезенка, лимфатические и гемолимфатические узлы и лимфоидные элементы в стенке различных органов и систем.
Развитие миелоидного кроветворения:
В развитии выделяют три периода:
Мезобластический
Гепатолиенальный
Медуллярный
Мезобластический (2 недель – 4 месяцев): первые клетки крови обнаруживаются у 13-19 суточного эмбриона в мезодерме желточного мешка. Интраваскулярно часть стволовых клеток крови дифференцируются в эритробласты (крупные клетки имеющие ядро). Экстраваскулярно образуются гранулоциты: нейтрофилы и эозинофилы. Активность мезобластического кроветворения снижается на 6 неделе и заканчивается на 4 месяце эмбриогенеза.
Гепатолиенальный (2 месяцев – 7 месяцев): в печени кроветворение начинается на 5-6 неделе, достигая максимума к 5 месяцу эмбриогенеза. Все форменные элементы - это эритроциты и тромбоциты в этот период образуются экстраваскулярно. К моменту рождения в печени могут сохраняться единичные очаги кроветворения. В селезенке очаги миелоидного кроветворения обнаруживаются с 20 недель эмбриогенеза, несколько позднее появляются очаги лимфоидного кроветворения, а с 8-го месяца эмбриогенеза в ней остается только лимфоидное кроветворение.
Медуллярный или костномозговой: начинается параллельно развитию костного скелета и продолжается всю жизнь. В полость первичной кость начинают врастать и дифференцироваться клетки двух типов: с 2-х месяцев механобласты (формируют ретикулярную ткань, которая заполняет все полости кости) и с 3-х месяцев - стволовые клетки крови, формируя островки гемопоэза. К 4-му месяцу эмбриогенеза ККМ становится главным органом кроветворения и заполняет полости плоских и трубчатых костей. У ребенка 7 лет ККМ в диафизах трубчатых костей бледнеет, появляется и начинает разрастаться желтый костный мозг. У взрослого человека ККМ сохраняется лишь в эпифизах трубчатых костей и в плоских костях. В старческом возрасте костный мозг (как красный, так и желтый) приобретает слизистую консистенцию и носит название желатинозный костный мозг.
Морфология красного костного мозга (ККМ)
Красный костный мозг (medulla ossium rubra ) – это центральный орган гемопоэза и иммуногенеза, содержащий популяцию стволовых клеток крови и участвующий в образовании клеток миелоцитарного и лимфоцитарного рядов.
Функции ККМ:
Гемопоэтическая - в красном костном мозге берут начало все ростки кроветворения на основе самоподдерживающейся популяции стволовой клетки крови
Иммунная - в красном костном мозге происходит антигеннезависимая дифференцировка лимфоцитов
Регуляторная – выделяющиеся в красном костном мозге гемопоэтины влияют на процессы кроветворения во всех органах гемопоэза, а синтезирующиеся цитокины регулируют иммуногенез.
У взрослого масса ККМ составляет 1,5 - 2 кг, это 4-5% от массы тела. Он имеет красный цвет и полужидкую консистенцию. Основу его или стромальный компонент образует ретикулярная ткань, состоящая из отросчатых ретикулярных клеток (ретикулоцитов) и межклеточного вещества, содержащего ретикулярные волокна. Она не только формирует трехмерную сеть, выполняя опорную функцию, но ее клетками синтезируются гемопоэтические факторы , без которых кроветворение не осуществляется. Ретикулоциты находящиеся вокруг стенки кровеносных синусов называют адвентициальными клетками. Эти клетки способны сокращаться, облегчая миграцию форменных элементов крови сквозь сосуды. Кроме ретикулоцитов стромальный компонент представлен адипоцитами , макрофагами, а также клетками эндоста (соединительнотканной выстилки костных полостей) - остеобластами и остеоцитами .
Адипоциты располагаются островками, обеспечивая энергией гемопоэз; заполняют объем, создавая давление, необходимое для функционирования синусов, а также вырабатывают БАВ, регулирующие объем кроветворной ткани.
Макрофаги выполняют различные функции: выделяют ростковые факторы и фагоцитируют клетки, «непрошедшие отбор»; макрофаги, мигрирующие из селезенки, приносят компоненты для синтеза гемоглобина, а костные макрофаги остеокласты – регулируют размеры костных лакун.
Остеобласты и остеоциты, формирующие жесткий остов поставляют микроэлементы необходимые для кроветворения.
Между ретикулярными клетками располагается большое количество полостей, в которых залегают гемокапилляры. Они очень тонкие и придают цвет ККМ. Вокруг кровеносных капилляров расположено множество свободно лежащих клеток крови миелоцитарного и лимфоцитарного рядов на разных этапах дифференцировки, а также самоподдерживающаяся популяция плюрипотентных стволовых клеток. Пролиферация в ККМ идет очень активно, создавая в сутки около 200 млрд. клеток.
Участки, где происходит пролиферация и дифференцировка клеток крови, получили название островков кроветворения. Эти островки, в общем,формируют гемопоэтический компонент.
Выделяют три типа островков:
Эритропоэтический островок содержит центрально расположенный макрофаг, называемый клеткой -нянькой, вокруг которого расположеныэритроидные клетки на разных стадиях развития (от колониеобразующей эритроидной клетки и эритробласта до ретикулоцита). Макрофаг выделяет ростковые факторы, с помощью сиалоадгезинов он удерживает вокруг себя эритроидные клетки, обеспечивая их железом за счет наличия в его цитоплазме трансферрина , связывающего 4 атома железа, также макрофаги вырабатывают эритропоэтин , витамин Д3 и фагоцитируют ядра, выбрасываемые из эритроцита в процессе созревания.
Гранулоцитопоэтические островки могут быть трех видов в зависимости от того какие гранулоциты образуются: нейтрофильные, эозинофильные или базофильные , чаще всего они локализуются вблизи эндоста. Каждый островок окружен слоем протеогликанов, что создает микроокружение для дифференцировки гранулоцитов. По мере созревания эта оболочка растворяется и гранулоциты, совершая амебовидные движения, мигрируют к синусам и уходят в кровоток.
Тромбоцитопоэтический островок локализуются у синусных капилляров, и включает мегакариоциты . Мегакариоциты - это очень крупные клетки с гигантскими дольчатыми ядрами. Он выдвигает ложноножку между эндотелиоцитами в полость капилляра и током крови эти участки отрываются, превращаясь в тромбоциты . Такой способ отрыва цитоплазмы называется «клазмотоз ». Из одного мегакариоцита формируются около 2 тыс. тромбоцитов.
Кроме этого в ККМ находится три категории лимфоидных клеток, лежащих вокруг сосудов:
Стволовые лимфоидные клетки, не имеющие рецепторов.
Предшественники Т – лимфоцитов , имеющие рецепторы и мигрирующие в тимус.
Предшественники В – лимфоцитов , в которых осуществляется уникальный процесс образования генов иммуноглобулинов.
Кроме этого в ККМ идет развитие моноцитов - будущих макрофагов.
Для того чтобы осуществились процессы гемопоэза и иммуногенеза нужны регуляторы, которые подразделяются на – стимуляторы и ингибиторы :
К стимуляторам относятся вещества, выделяемые клетками стромы, гормоны, синтезируемые в других органах: эритропоэтины в почках, легких, печени; тироксин щитовидной железы, соматотропный гормон гипофиза.
К ингибиторам относятся вещества, вырабатываемые зрелыми форменными элементами крови по принципу обратной связи (кейлоны), тканевые гормоны – интерферон, простагландины, гормоны коры надпочечников.
Сосудистый компонент ККМ представлен мощной сосудистой сетью, имеющей особенности в строении. Существуют 2 способа кровоснабжения костного мозга.





